Chapter 5: Rationale of this Study
5.1 Limitations of Previous Studies
The evidence reviewed in the previous chapter shows that neuroanatomical variability correlates with behavioral differences between species, within other species (specifically rats), as well as within clinical and normal human populations. This evidence is consistent with the hypothesis that neuroanatomical variation causes differences in behavioral ability. Nevertheless, a number of important questions remain to be addressed.
First, the causality of the correlations has not been demonstrated. In order for the associations between neuroanatomical and cognitive dimensions to be evolutionarily relevant, they must be shown to be causally connected. That is, the neuroanatomical variability must be shown to actually cause differences in cognitive ability, not just be associated with it. It is possible for neuroanatomy and cognitive ability to be correlated (through the intermediate influence of other variables) without one directly causing the other. The extent to which the associations reported in humans are not directly causal is not presently known.
Second, the studies on human subjects have generally limited the range of cognitive tasks that have been investigated, for understandable pragmatic reasons. For the most part the emphasis has been on IQ-type tests. This is reasonable given the nature of IQ. Cognitive test scores on
diverse batteries of tests are generally positively correlated, with the number and size of the positive correlations among such a battery of tests far exceeding that expected by chance (Jensen 1987). This fact forms the theoretical justification for the concept of a single ability factor, called g , underlying a large portion of the variability among individuals on a set of cognitive tests (IQ test scores are estimates of g). Thus, it is understandable that the initial focus of brain size / behavioral ability studies would be on IQ. Nevertheless, it is quite possible that other, more specific cognitive dimensions were as important, or more so, than g during human evolution. Potentially important cognitive dimensions can be gleaned from an understanding of the behavioral changes that occurred during human evolution (as outlined in chapter 3) and also from a consideration of the neuroanatomical areas that underwent the greatest change. Such dimensions include: aspects of linguistic processing (including syntax and access to the semantic meaning of a given symbol), spatial processing, and dimensions of prefrontal processing.
It should be noted here that the existence of a correlation between some cognitive dimension and an evolutionarily important neuroanatomical variable does not constitute evidence that cognitive dimension was specifically selected for. It is possible that it is a side effect of selection on another variable. Furthermore, if we find that cognitive dimension "A" correlates more strongly with, e.g., brain size than dimension "B", this does not constitute evidence that "A" was selected for more strongly than "B". It could be that "B" was selected for more strongly, but that the resulting neuroanatomical changes inadvertently had greater effects on "A". Nevertheless, the relative differences in strength of association are not irrelevant to our understanding of human cognitive evolution. Clearly, if "A" correlates with brain size while "B" does not, it would be hard to maintain that selection for "B" was the driving force behind brain size evolution, and that "A" was irrelevant.
Third, the neuroanatomical focus has been on overall brain size. There are other interesting neuroanatomical components which might have greater correlations with various cognitive dimensions than overall brain size. An obvious possibility here is prefrontal cortex, as explained in chapter 4. Other possibilities include the cerebrum (which excludes the brain stem), cerebellum, grey vs. white matter, and so forth.
Fourth, it is not known whether or not the costs of brain size, which are apparent in cross-species analyses, can be shown to exist within humans. If the costs are high within humans (as they are between-species) then we should expect the payoffs to be high within humans as well. Conversely, if the costs are low within humans, the payoffs will likely be correspondingly low.
Finally, previous MRI studies obtained brain images with a spatial resolution no finer than 5 mm3 (because of individual slice thickness). In addition, most scanning routines left gaps of non-imaged brain tissue between each slice, increasing the effective resolution to no less than 7 mm3.
5.2 Contribution of the Present Research
5.2.1 Within-Family Correlations as a Test of Causality
If any brain/behavior relationships are to be meaningful from an evolutionary perspective they must be directly causally linked: a change in brain anatomy must have some influence on behavior. The argument that brain size has a direct causal influence on IQ is entirely reasonable given what we know about brain, behavior, and evolution. However, the demonstration of a correlation between two variables does not prove that one variable has a direct causal influence on the other, even though it is an important first step. A correlation between two variables is a necessary consequence of a direct causal relationship, but because other effects can lead to an association between variables, it cannot constitute proof that one variable is causing change in the other.
One way in which a correlation can exist between two variables without there being a direct causal influence is when both are independently influenced by a third variable (or set of variables). Such an association will be referred to here as a non-intrinsic correlation (following Jensen 1980). An example of a non-intrinsic correlation is that between weekly soft-drink sales and the incidence of polio (before introduction of the Salk vaccine), which occurred because polio epidemics were most severe during the summer, not because soft-drinks were causing polio (Freedman et al. 1978). With respect to brain/behavior associations, it is possible that some environmental influence is acting on both variables independently. One such influence might be family income (or some measure of social class or socio-economic status) which independently affects diet, health care, and other variables that affect the growth of the body, while at the same time influencing the quality of education available to individuals and consequently their IQ test scores as well. Figure 5.1 illustrates the possible web of causal associations involving socio-economic status (SES) which could lead to a non-intrinsic association between brain size and cognitive test performance. The reciprocal arrows from cognitive test performance to SES serve to illustrate the possibility that test scores might have a causal effect on SES. This could occur via the long-term economic benefits of doing well on these sorts of cognitive tests, which could lead to better nutrition, health care, and other possible correlates of somatic growth, thereby leading to larger brain size. But whatever the true direction of these influences, SES could at least theoretically explain the
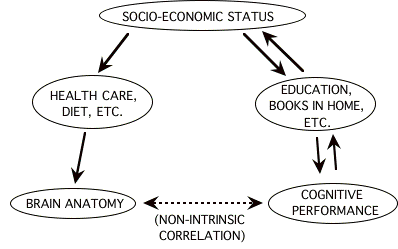
"Figure 5.1 Hypothetical causal links between SES, brain anatomy, and behavior" \l 1 Figure 5.1: Hypothetical causal associations involving socio-economic status which could lead to a non-intrinsic correlation between brain anatomy and cognitive performance. A non-intrinsic correlation is one in which no direct causal effect occurs between the variables of interest.
existence of a non-intrinsic association between brain anatomy and cognitive test performance.
Another factor that could result in non-intrinsic correlations is cross-assortative mating. This occurs whenever there is a non-random association between two variables across mating pairs: i.e., when variable "A" in one parent is correlated with variable "B" in the other, and vice-versa. These variables would then be correlated in the offspring (and therefore the population as a whole), but not because variable "A" is causing differences in variable "B". They would be correlated because the genes influencing variable "A" (derived from one parent) would be more likely than chance to find themselves in bodies that also have genes influencing variable "B" (derived from the other parent). This could also work if the influences are environmental, instead of (or in addition to) genetic ones.[1] Note here that conscious knowledge among mating pairs of any cross-correlation on a particular set of traits is not necessary for the association to occur. Also, it is known that simple assortative mating (in which variable "A" is correlated across mating pairs, without regard to other variables) exists for many traits, both anatomical and (especially) behavioral (the evidence for this will be reviewed below).
Now it is entirely possible (and in fact quite likely) that at least some portion of the association between brain size and IQ is the result of such non-intrinsic influences: either the influence of third variables like SES, or through some form of cross-assortative mating. The crucial question, from an evolutionary perspective, is how much of the association is attributable to these non-intrinsic influences? That is, a clear understanding of human brain evolution requires that we know the size of the direct causal influence that brain anatomy has on behavior. For example, note that the relative adaptive benefit of increasing brain size over our evolutionary history is proportional to both: 1) the direct causal association between brain size and behavioral ability, and 2) the ratio of costs-to-benefits of increasing brain size. If the direct causal influence of brain size on behavior is very small, then this will influence our judgment about either the relative adaptive benefit or the ratio of cost-to-benefits of increasing brain size. Thus, the size of the direct causal association between brain size and behavior is an important piece of the puzzle of human brain evolution.
How, then, can the size of the direct, causal association between brain anatomy and behavior be estimated? There are in principle several ways in which this can be investigated. Perhaps the best way is to compare the correlation between brain anatomy and behavior in pairs of identical twins reared in uncorrelated environments (e.g., adopted into different families). In this situation, the cross-correlation between one twin's brain anatomy and the other's behavior is a direct estimate of the true genetic contribution to the phenotypic resemblance between these variables (Plomin et al. 1990). Unfortunately, identical twins reared in separate homes are very rare, and obtaining access to twin registries is difficult. However, there are other genetically informative designs that could be used which vary in their complexity and difficulty.
A reasonable place to begin addressing this question is through a within-family design, in which pairs of siblings are compared on the traits in question. Such a design allows one to estimate both between-family and within-family correlations (Jensen 1980, 1989, 1990). Between-family correlations are calculated from mean family values on the variables in question. Within-family correlations are derived from the signed sibling differences for the variables in question (see the next chapter for more detail). Between-family correlations include covariance that is due to major (between-family) environmental influences (like SES) as well as that due to cross-assortative mating. Within-family correlations, on the other hand, by definition do not contain any covariance due to between-family influences. They also do not contain any covariance due to cross-assortative mating, due to the specifics of meiosis. Genes among siblings within families are a random assortment of their parents genes. Thus, even if there were any cross-assortative mating, thereby leading to a statistical association among genes for two traits in the overall population, these genes would be randomly assorted with respect to each other among siblings (and therefore no association between the variables) within families.
The statistical independence of between- and within-family correlations can be visualized by comparing the two graphs in figure 5.2, which represent hypothetical associations between two variables for 6 individuals in 3 families (each family pair is connected by a separate line). In figure 5.2a, the two variables are correlated positively both between-families and within-families. In figure 5.2b, by contrast, the two variables are correlated positively between-families (i.e., the family means for the two variables are perfectly correlated), but they are negatively correlated within families (the sibling with a higher value on the x-axis is always lower than the other sibling on the y-axis). Thus, there is no necessary association between the two sets of correlations.
The basic argument is that if differences in brain anatomy actually cause differences in a particular behavior, then offspring who differ from

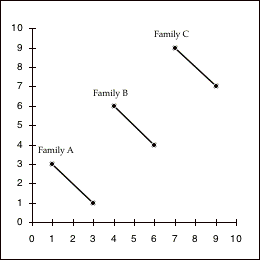
Figure 5.2: The theoretical independence of between versus within family associations. Figure A shows a hypothetical set of data in which the between and within family correlations are identical. Figure B shows a situation in which the between family correlation is positive, even though the within family correlation is strongly negative.
their siblings neuroanatomically should also differ (above chance) in a manner consistent with the between-family association for that behavior . This is because siblings (unless they are identical twins) are not born with identical genetic influences. Thus, if they differ neuroanatomically, they must also differ behaviorally if there truly is a genetic association between brain and behavior. Furthermore, because the within-family correlation is not inflated by between-family environmental effects like SES, or by cross-assortative mating, it is a better estimate of the direct genetic association between two variables, and thus is more relevant for an evolutionary understanding of brain/behavior relationships.
There is one caveat, however. If the within-family environmental influences work in opposite ways on the two variables of interest, and if they are of sufficient magnitude, then the within-family correlations between the two variables could be zero even if the genetic correlation between them is not. This would require, e.g., that the within-family environmental influences that have the effect of increasing brain size, must, at the same time have the effect of decreasing IQ (or whatever cognitive dimension correlates with brain size between families). While this may be possible, it is hard to imagine, a priori, what sorts of influences would have such effects.
In any case, the results of a within-family design will be a crucial first step in unraveling the genetic and environmental influences underlying brain/behavior relationships. If the correlations are similar both between and within families, this would support the hypothesis that these dimensions of brain anatomy and behavior are influenced by the same genes, and/or that the environmentally induced variation in brain anatomy also has behavioral effects. If the correlations differ significantly, it would call into question the validity of the correlations that have been reported between, e.g., brain size and behavior (as reviewed in chapter 4). Thus important knowledge will be gained in either case.
5.2.1.1 Height/IQ and Myopia/IQ Associations: Test Cases
Examples of this sort of analysis can be found in studies of the relationship between various dimensions of body size and IQ, as well as between myopia and IQ. Correlations have been demonstrated between height and IQ test scores (and also between height and income; see Jensen 1990 for a review). These associations are typically smaller than those found in MRI-derived brain/behavior studies, though they are still statistically significant. Nevertheless, they raise the general question of why gross biological dimensions like height would correlate with behavioral ones at all. Note that this is not the case with brain size and IQ studies, for which there is a theoretical justification for expecting some sort of an association.
Interestingly, several studies have shown that the correlation between height and IQ is actually zero within families: taller siblings are not more likely to score higher on IQ tests than shorter siblings (Laycock and Caylor 1964, HusŽn 1959, Jensen 1980, Nagoshi and Johnson 1987). Thus, as explained above, the association that is found between families (and in the population at large) is not likely due to a direct causal influence between height and IQ (either IQ causing differences in height, or vice versa). It is not known whether the association between height and IQ is due to an environmental explanation, to cross-assortative mating (the explanation favored by Jensen 1990), or some combination, but it does suggest that the same question should be asked of brain/behavior associations.
It is also known that myopia is associated with higher IQ (see Cohn et al. 1988 for a review). Empirically, it does not appear that this association is caused by a greater degree of "nearwork" (e.g., reading) in higher IQ subjects. Furthermore, behavior genetic studies point to a clear genetic influence on variance in ocular refraction (Cohn et al. 1988). In contrast to the situation for height and IQ, the myopia/IQ association is also found within families. Cohn et al. (1988) showed that higher IQ siblings also tend to have a greater degree of myopia, even though the lower IQ siblings tended to spend more time studying and read more books on average. A pleiotropic relationship, in which genetic influences on IQ also influence the incidence of myopia, would appear to be the most likely explanation in this case.
Thus, within family studies have provided important clues to the causal nature of associations between biological and behavioral variation. If brain anatomy has a direct causal influence on behavior (i.e., where brain size differences directly affect behavioral differences), then either there must exist a demonstrable within-family association, or else we must be able to show that within-family environmental influences have opposite effects on brain and behavior.
5.2.1.2 Brain size/IQ Studies of Between- versus Within-family Correlations
None of the MRI-based studies of brain size and behavior reviewed in chapter 4 have addressed this issue. However, two studies have addressed it with respect to head-circumference/IQ correlations. Jensen (1994) analyzed monozygotic (MZ) and dizygotic (DZ) twins and found that the within-family correlations between head-circumference and g factor scores (an estimate of IQ calculated from the common variance in a diverse set of cognitive tests) was significant for DZ twins (r= 0.28, p< 0.05) but not for MZ twins (r= 0.01, NS). This is consistent with the hypothesis of a causal biological association between these variables because the differences between MZ twins within-families are by definition entirely due to environmental differences. However, he found essentially no correlation between-families for DZ twins (r= 0.04, NS, though the correlation between-families for MZ twins was significant; r= 0.31, p< 0.05). Further troubling was the finding that the multiple correlation of head length, width, and circumference with g scores within-families was approximately the same for both MZ and DZ twins (MZ r= 0.21, DZ r= 0.28; both NS), even though the hypothesis predicts that the correlations should be significantly different. In a different analysis of a large body of data from the National Collaborative Perinatal Project (total N= 28,992), Jensen and Johnson (1994) found that the within-family correlations between head circumference and IQ in 7 year olds were almost half as large (57%) as the corresponding between-family correlations (mean within-family r= 0.12, p<0.05; mean between-family r= 0.21, p< 0.001).
These two studies suggest that a significant portion of the correlation between head circumference and IQ is not attributable to direct causal connections between the two variables. Given that MRI-based studies consistently report larger correlations to IQ than do head circumference studies, I sought to investigate the same question of between-family vs. within-family correlations using MRI-derived brain size estimates. Since the within-family correlation is potentially the most meaningful from the standpoint of explaining brain size evolution, it is crucial that this be estimated as accurately as possible. Thus, the present study uses a within-family design, and within-family associations were estimated for all brain/behavior associations that were found.
5.2.2 Expanding the Variety of Cognitive Dimensions Addressed
A wide variety of cognitive domains were addressed in the present research. Psychological studies of individual differences (psychometrics) has delimited four major cognitive factors (in addition to many lesser ones; Jensen 1987). These factors are named according to the types of tests that load (correlate) most highly with each factor. The verbal factor is so described because the tests which load most highly on it all share some explicit linguistic component. Examples include vocabulary, reading comprehension, and related tests. The spatial factor is defined by tests which lack obvious linguistic components, but instead deal with the location, interrelation, and/or movement of objects in space. Examples are tests of how three-dimensional objects might look if rotated into different views ("mental rotation" tests). As noted in chapter 3, spatial ability is not as cleanly described by single tests, unlike the verbal factor (Lohman 1988). A third factor involves explicit tests of short-term memory. Finally, there is a general factor, designated g , which cuts across these categories.
As noted above, cognitive tests almost without exception correlate positively with each other, even if they differ substantially in the sorts of abilities they require (Jensen 1987). For example, the vocabulary and block design (spatial) portions of the widely used Wechsler Adult Intelligence Scale (WAIS) correlated on average r= 0.52 across all age groups in their standardization sample (N = 1880), even though these two tests appear to require completely different abilities (block design has no obvious verbal component, and vocabulary has no spatial component; Wechsler 1981). G typically explains more than 40 percent of the total variance in varied sets of cognitive tests, while the other factors combined account for only about 15 percent (the remainder being variance specific to individual tests; Jensen 1987). It is important to note, however, that these test batteries typically do not contain tests known to be specific to prefrontal functioning, which generally correlate very little with g -loaded tests.
There are practical limitations for any given study in the number of cognitive dimensions that can be investigated. At the same time, since an important contribution of the present study is to broaden the range of behavioral features investigated, it is important to include tests of the four basic factors (outlined above), so that the variance in any new tests (which might be shared with these basic dimensions) can be controlled for. If, for example, we find that some new test of interest is correlated with brain size, we cannot be sure that this correlation is due to some unique aspect of the demands of this new test unless we first determine the extent to which it correlates with g or one of the other major cognitive factors. Only if we find that the correlation with this new test is not due to its correlation with these other more general factors can we be confident that we have discovered a unique correlate of brain size (and/or any other neuroanatomical dimension of interest). For this reason, the following tests were included as markers for the basic factors (detailed descriptions of the tests appear in the Methods chapter). As an estimate of g, a adaptation of Raven's Progressive Matrices was used (This is widely acknowledged to be an excellent measure of g; Jensen 1987). A standard vocabulary test, included in the Multidimensional Aptitude Battery (Jackson 1984) was used as a marker for the verbal component. A computerized adaptation of the Vandenberg and Kuse (1978) Mental Rotation Test, in a reaction-time format, was used to gauge spatial abilities. No test of short-term memory was included, because it was deemed less likely than other dimensions (see below) to have contributed significantly to brain size (this was a pragmatic decision; it is possible that this was a tactical mistake that will need to be addressed in future studies).
5.2.2.1 Prefrontal tests
In addition to the basic dimensions, tests that have shown some sort of specificity to prefrontal cortex were included for the first time in any study to date. As has been discussed in previous chapters, the prefrontal cortex has undergone much greater change than any other region during our evolutionary history. Unfortunately, because of the types of behavioral deficits evidenced by individuals with prefrontal damage, it has proven difficult to create reliable and valid tests of prefrontal function that are both easy to administer and (most importantly) show some sort of gradation of ability across normal subjects. It is known that subjects with prefrontal damage often score normally on standard IQ tests while showing gross problems on everyday tasks involving some component of planning (Shallice and Burgess 1991). As a starting point, it was decided to include four tests which generally show prefrontal specificity (e.g., in brain damaged patients) while at the same time demonstrating significant individual variation in normal subjects (Shallice and Burgess 1991, Daigneault et al. 1992, Lezak 1983). The four tests selected included: The Trails test (Reitan and Wolfson 1985), the Wisconsin Card Sort Test (WCST, Heaton 1981), the Verbal Fluency test (Thurstone and Thurstone 1941), and the Stroop test (Stroop 1935; Golden 1978).
The trails test is essentially a connect-the-dots task with two parts. The first (part A) involves only numbers, arranged such that the subject can obtain information on where they will have to go after the immediate goal is reached. For example, the number 14 is located very close to the path between 12 and 13, such that the subject can potentially plan on the way to 13 which direction they will have to go next. The second portion of the Trails Test (part B) involves alternating between numbers and letters of the alphabet, such that the subject must connect the dots in the following pattern: 1 -> A -> 2 -> B -> 3 -> C ... etc.. These are also arranged in the same manner as in part A, such that the subject can plan ahead.
The WCST is a test which requires the subject to sort a long series of cards according to certain criteria which keep changing. The cards are designed such that there are three basic, independent ways the cards could be sorted: either by shape of the objects on the card, by number of objects, or by the kind of pattern filling the objects. The subject must first figure out which of these potential sorting criteria is actually being rewarded, simply on the basis of being told their choice (which is initially a guess) is "correct" or "incorrect". After the subject demonstrates they have figured out which criteria is currently being rewarded, the correct criteria is changed and the subject must figure out (on the basis of simple "correct" or "incorrect" feedback responses) what the new sorting criterion is. This test is not so much a test of planning, but rather a test of how flexible the subject is to changing criteria.
The Verbal Fluency task requires the subject to list as many words as possible that start with a particular letter of the alphabet. Three different letters are used (to minimize the impact of "blanking out" on any one letter), and a one minute time limit is used for each letter. Since words are not stored in our mental lexicon alphabetically, this is as much a test of the ability to suppress obvious associations as it is a test of vocabulary size. For example, the word "chair" for most people brings to mind the word "table" or "sitting," both of which are incorrect responses and must be quickly suppressed to obtain the best scores.
Lastly, the Stroop test presents the subject with conflicting cues: linguistic and visual (color), and requires that the subject pay attention only to the color cue. For example, the word "red" is written in blue ink, and the subject must respond "blue". This is very difficult for most subjects (including normal ones). It would appear to be essentially a test of the ability to suppress obvious linguistic cues. There are control tasks which allow for differences in simple reading and color identifying ability (the reading ability task could also potentially be used to assess phonological production speed).
From these descriptions, it is clear that none of these tests can be considered a pure test of prefrontal function. It is essential to include a basic battery tapping verbal, spatial, and general (g) cognitive abilities, such that these other dimensions may be factored out of the prefrontal tests. As a basic example, Daigneault et al. (1992) conclude that there are (weak) prefrontal factors which can be extracted from a set of prefrontal tests. However, since their test battery does not include markers for g, verbal, spatial, or memory, it is quite possible that these factors are not specific to prefrontal functioning at all, but instead are a further manifestation of these other general cognitive domains.
5.2.2.2 Additional linguistic tests
In addition to a basic vocabulary test, several other dimensions of linguistic processing were investigated. Three dimensions were of particular interest: phonological processing, semantic retrieval and syntax processing. Phonological processing is not essential to language, as is evidenced by the existence of various sign languages, but semantic retrieval and syntax clearly are essential. With respect to syntax, however, linguists in the generative grammar school expressly deny that there are meaningful individual differences in syntax and grammatical ability among individuals.[2] Similarly, the evolutionary psychologists Tooby and Cosmides (1990) hold that basic adaptations of a species (including cognitive adaptations) will be essentially invariable. They argue that the network of interlocking parts that make up a whole organism is so complex that anything more than minor tinkering would cause problems when different parts are reshuffled during genetic recombination. However, there must have been meaningful individual variation at each point along the continuum of populations leading from the language-less ancestors to the language-using descendent populations. It is exactly this variation which we are concerned about in this study.
There are a large number of cognitive tests which tap different linguistic processes. Phonological processing speed was gauged by a portion of the stroop test discussed above. The first part of this test requires the subject to simply read a random ordering of three color words: red, green and blue (this is used a control task for the stroop condition in which the color and word do not match; see method section below). Unfortunately, there were no existing tests which allowed such components as syntax and semantic retrieval to be studied in isolation from other components. For this reason, two tasks were invented for this study specifically to address these two linguistic processes.
Semantic retrieval was studied through a reaction-time task which required the subject to correctly match up a word to a picture (which might or might not correspond). The time it took to respond correctly is a measure of how fast the individual could 1) decode the meaning of the word, 2) process the picture, 3) compare the results, and 4) respond. In order to control for differences in vocabulary size, only a few simple words were used which every English speaker could be expected to have highly over-learned: circle, square, triangle, and star (see the next chapter for details). Because of the need to limit the testing time (out of concern for test fatigue on the part of the subjects) as well as the desire to give as broad a set of cognitive tasks as possible, only these four words were used. In order to control for individual differences in the ability to simply respond quickly, irrespective of the task, a simple reaction time task was also given (on the same computer apparatus). It is important to recognize that no single task will be perfect, but that this test would certainly give some indication of individual differences in semantic retrieval.
Syntax processing was tested by taking advantage of the fact that not all syntactic structures are as easy to process as others, even for fluent speakers. A well known example occurs when relative clauses are embedded within other clauses (Yngve 1960, Miller and Chomsky 1963, Foss and Lynch 1969, Bever 1970, Kuno 1974, Cook 1975, Abe et al. 1988). For example, compare the two following sentences:
1) The man kissed the woman who ran.
2) The woman whom the man kissed, ran.
In sentence 2, the clause "...whom the man kissed..." is embedded in the center of the clause "The woman...ran". This type of syntax can be referred to as "center-embedded". In sentence 1, the clause "...who ran" is added to the right of the clause "The man kissed the woman," and can therefore be referred to as "right-branching". Center-embedded sentences are normally harder to process than right-branching ones. Furthermore, it turns out that there are individual differences in how difficult center-embedded sentences are to process (Cook 1975). This fact was used as the basis for a test for which it is possible to extract a measure of syntax processing that is independent of all other aspects of linguistic processing.
The basic format of the test followed that used in the seminal studies on sentence verification by Clark and Chase (1972), in which the subject was presented with simple sentences which might or might not correspond to pictures presented at the same time. Their data showed that the time required to verify that the sentence and picture corresponded was related in a regular way to the linguistic complexity of the sentence (see also Carpenter and Just 1975). However, in these studies the semantic aspects of the sentence are intertwined with the syntactic aspects, such that the differences due solely to syntax can not be disentangled.[3] In the present study, this was controlled for by comparing reaction times for pairs of sentences which differ in clause structure (center-embedded versus right branching), but are identical in the semantic units used. For example, compare sentences 3 and 4 below:
3) The square is beside the circle which is above the star.
4) The square which the circle is beside is above the star.
These two sentences have exactly the same number of words, and the words used are exactly the same. The only difference between these sentences is in their syntactic structure. Thus, differences in mean reaction time between sets of sentences like these cannot be due to semantic differences.
Unfortunately, it is possible that the differences between individuals on how much these syntactic structures influence sentence verification times are due to differences in the richness of their past linguistic experience. If some individuals are more likely to have heard center-embedded sentences before than others, this past experience might result in smaller differences in verification times for these individuals because of this. While the experimental design used here cannot exclude this possibility, at least two correlates should appear in the pattern of results. First, individuals who have a greater depth of linguistic experience will likely do better on both sentence types. That is, we might expect a correlation between 1) mean reaction time for both sentence types and 2) the difference between center-embedded and right-branching sentences. Secondly, we should expect the difference between the sentence types to correlate with the degree of educational background or socio-economic status. Both of these correlates can be checked on the present data (see below).
Again, no test is perfect, but these tests specifically address the basic linguistic dimensions of syntax and semantic retrieval.
5.2.2.3 Throwing accuracy
Throwing accuracy has not been measured in any MRI study to date. In this study it was measured in a dart throwing task in which the subject was given 12 throws to get as close as possible to a central spot located on a cork board 275 cm away.
5.2.2.4 Social Dimensions
There are a few pencil and paper tests that investigate social intelligence (e.g., Marlowe 1986), but it seemed that the most direct way to investigate social ability would be to ask questions relating specifically to the number of social contacts of each subject. As discussed in chapter 3, there is extensive evidence that relative neocortex size is highly correlated across primate species with the size of the social group. Thus, the most direct comparison within humans would be some measure of the number of social contacts reported by each individual subject. The subjects were asked to count (in front of the experimenter) the number of individuals listed in their address books (and to subtotal the number that were relatives). They were asked to estimate the number of individuals they talked to for more than five minutes in the prior week, as well as the number of individuals they spent time with socially in the average month. Lastly, they were asked to rate themselves as to how social they considered themselves, and to compare their own degree of sociality to that of their participating sibling (pairs of siblings were tested - see below). Again, none of these can be considered perfect measures of social ability or social intelligence. Nevertheless, it represents a crucial initial step in addressing this fundamental question.
5.2.3 Expanding the Neuroanatomical Subdivisions Addressed
There are a number of neuroanatomical subdivisions which are of interest in addition to overall brain size. The cerebral cortex is the site of conscious awareness, processing of primary sensory information, cross-modal associations, spatial processing, language (in humans), planning, and other complex mental functions. The lower brain stem, including the medulla, is involved in basic autonomic functions, such as breathing, heart rate, etc.. The cerebellum appears to be primarily involved in (sub-consciously) fine-tuning muscle control and tone, although there are some indications that it may also contribute to higher cortical functions such as language (Leiner et al. 1986). Within each of these major divisions, neural tissue can be classified into two types: grey and white. Neuronal cell bodies are found in grey matter, while axons (which carry signals across relatively large distances) are found primarily in white matter. As a rough approximation, white matter can be thought of as connective wiring between processing units (which are areas in the cerebral cortex and sub-cortical nuclei), while the grey matter represents the processing units themselves. The axons in white matter are insulated with layers of myelin - a fatty substance which appears white on visual inspection. The white and grey regions can be quite distinct on MRI scans, given the correct scanning parameters, which allows accurate quantification of these two variables.
5.2.4 What is the Effect of Increasing Brain Size on Maturation-Rate?
As was pointed out in chapter 3, one of the most obvious evolutionary costs to large brain mass appears to be that it takes an increasingly long time for larger brains to mature. The correlation between age at first breeding in females and overall adult brain size is r= 0.96 across a broad sample of 47 primates (Harvey and Clutton-Brock 1985). The longer it takes offspring to mature, the fewer offspring an individual can produce per unit time. Thus, assuming the correlation is causal, individuals with smaller brains should have an evolutionary advantage over those with larger brains, all else being equal.
The extent to which brain size correlates with rate of maturation within any species has apparently not been addressed to date. By recruiting women, it is possible to estimate rate of maturation by asking the subjects to recall the age at which they had their first menstrual period. Recalled age at menarche has been shown to be a reliable and valid measure, even over long periods of time (Geary 1989, Damon and Bajema 1974, Bean et al. 1979). However, the extent to which variation in age at menarche can be ascribed to genetic differences does not appear to be large. Johnston (1974) reviews evidence suggesting that at least 10 to 15 percent of the variation in age at menarche is genetically influenced. Tanner (1962, cited in Johnston 1974) showed that monozygotic twins differed by only two to three months in age at menarche, compared to 12 months difference on average for dizygotic (fraternal) twins. Socio-economic status does not appear to have a significant effect across the board, though the number of sibs in a family does (independent of socio-economic status; Johnston 1974).
Since the present study will also be comparing sibling differences, major between-family environmental effects (such as socio-economic status, number of siblings, etc.) will be controlled for, with the hope that intrinsic genetic influences will therefore be emphasized. This is probably the best we can do within humans. Obviously a large sample size would be nice, but not practical given the cost of acquiring MRI scans on each subject. Thus, this question is not the focus of the present study.
5.2.5 Additional Variables of Interest
Because correlations could occur between neuroanatomical and cognitive dimensions for reasons other than direct causation (e.g., through the influence of a third variable, as pointed out above), the possible effect of the most obvious such candidate, socio-economic status (SES), will be addressed as well. Four pieces of data will be used to estimate this parameter: 1) mother's years in school, 2) father's years in school, 3) socio-economic status rating of parents job (using a standard SES index; see methods section), and 4) a questionnaire, known as the Home Index of status items, which asks about items and aspects of the subject's home environment which are correlated with SES (Gough 1971).
5.2.6 Increasing MRI Spatial Resolution
Technical advances are occuring constantly in MRI technology, such that it is now possible to acquire brain image data with a spatial resolution of 1.3 mm3 (0.9375 mm X 0.9375 mm X 1.5 mm), with no gaps, covering the entire brain, and providing excellent grey/white tissue differentiation. This represents an almost 4-fold improvement in spatial resolution over the highest resolution MRI study yet published.
5.3 Summary of Research Goals:
The important questions addressed by this research include:
1) Are there any correlation between neuroanatomical variability and behavioral ability within families (which is the only correlation that really matters in an evolutionary sense)?
2) Are there any significant differences in strength of correlations among different behavioral measures (e.g., spatial, verbal, prefrontal, degree of sociality, throwing accuracy, etc.) with brain size or any other major neuroanatomical divisions?
3) Are there major subcomponents of the brain that are more highly related to these behavioral differences than others?
4) Does rate-of-maturation vary significantly with brain size or any other neuroanatomical component?